




|
22/09/2023 Lượt xem: 542
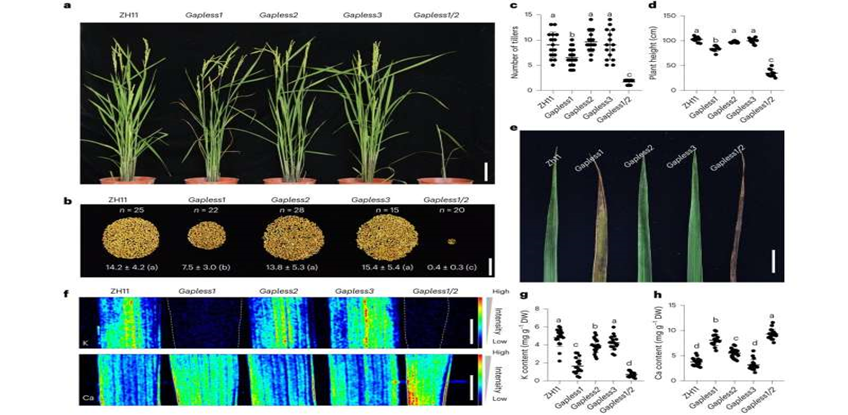
Protein GAPLESS cần thiết cho sự sinh trưởng, năng suất và cân bằng dinh dưỡng của cây lúa Hình a: Hình ảnh đại diện của ZH11 và những đột gapless biến khe hở khác nhau được trồng trên ruộng lúa ở giai đoạn đầu. Các đột biến gapless1, gapless2, gapless3 và gapless1/2 là các dòng của gapless1 – 1; gapless2- 1 và gapless3 -1 và gapless1-1/2 – 1. Ảnh được chụp sau khi cây được đào lên khỏi ruộng lúa. Hình b: là hình ảnh đại diện của hạt từ một cây có kiểu gen khác nhau. Thống kê trọng lượng hạt trên mỗi cây được đánh dấu. Hình c và d: Xác định số bông và chiều cao cây của ZH11 và những đột biến hở gapless khác nhau. Số liệu phân tích trên các mẫu độc lập. Biểu đồ c; n= 15 đối với ZH11 và gapless3. N= 21 trên gapless1, n= 11 đối với mẫu gapless2 và gapless1/2; n= 13 trên mẫu gapless3. Hình e: hình ảnh đại diện cho các lá già từ ZH11 và những đột biến gapless khác nhau trên cùng ruộng lúa. Hình f: Hình ảnh chiếu huỳnh quang tia X cho thấy sự phân bbos kali và canxi trong lá của các kiểu gen khác nhau. Biểu đồ g: nồng độ kali; h: nồng độ canxi trong lá già của ZH11 và gapless1/2. n = 17 đối với mẫu gapless1 và gapless3; n= 20 đối với mẫu gapless2 Chữ cái trong đồ thị đo 10cm (a), 5 cm (b), 2cm (e) and 5mm (f) là số liệu đại diện cho giá trị trung bình ± s.d. các chữ cái khác nhau c,d,g và h biểu thị sự khác biệt đáng kể ở P< 0,05 ( phân tích ANOVA một chiều của Tukey. Nguồn: Nature Plants (2023). DOI: 10.1038/s41477-023-01503-z. Trong một nghiên cứu được công bố trên tạp chí Nature Plants, một nhóm nghiên cứu do giáo sư Chao Daiyin từ Trung tâm chuyên nghiên cứu Thực vật Phân tử của Viện Hàn lâm Khoa học Trung Quốc dẫn đầu đã xác định được một nhóm protein mới trong thực vật có tên là GAPLESS và tiết lộ rằng các protein trong nhóm này làm trung gian cho sự bám dính giữa thành tế bào và màng sinh chất ở đai Caspari (CS), một rào cản trong nội bì rễ. Độ bám dính này giữ vai trò quan trọng trong việc kiểm soát sự vận chuyển chất dinh dưỡng và sinh trưởng của cây lúa. Các sinh vật sống cần có hàng rào để kiểm soát việc vận chuyển và trao đổi chất. Ở động vật, các tế bào biểu mô ruột được neo chặt thông qua một loạt các protein tổng hợp màng tế bào, tạo thành các "mối nối chặt chẽ" để bịt kín khoảng trống giữa các tế bào nhằm kiểm soát việc hấp thụ chất dinh dưỡng đồng thời ngăn chặn sự rò rỉ của chúng. Rễ là một cơ quan của thực vật có chức năng hấp thụ và vận chuyển nước và chất dinh dưỡng, có cấu trúc và chức năng tương tự như ruột động vật. Tế bào nội bì rễ, giống như tế bào biểu mô ruột, có chức năng kiểm soát sự vận chuyển nước/chất dinh dưỡng và ngăn ngừa rò rỉ nước/chất dinh dưỡng. Tuy nhiên, do có sự hiện diện thành tế bào nên các tế bào nội bì của rễ cây không thể thiết lập được các mối nối chặt chẽ trực tiếp như tế bào biểu mô ruột. Ở thực vật đã phát triển một cấu trúc thành tế bào độc đáo gọi là đai Caspari (CS) nhằm ngăn chặn sự khuếch tán tự do của nước và chất dinh dưỡng. Được đặt theo tên người đã phát hiện ra nó là Robert Caspary, cấu trúc này bao gồm gỗ hoá không thấm nước, ngăn cản sự khuếch tán tự do của các chất hòa tan trong nước. Năm 1935, nhà khoa học người Mỹ Bryant đã phát hiện ra màng sinh chất tiếp theo là đai Caspari (CS), còn được gọi là miền CS (CSD), bám chắc vào đai Caspari, giống như các mối nối chặt chẽ ở động vật. Sự kết dính này cực kỳ mạnh mẽ và thậm chí không thể phân tách bằng các phương pháp xử lý thẩm thấu cao có khả năng phân tách riêng các tế bào. Sự bám dính này được cho là đóng một vai trò quan trọng ở thực vật. Do khó nghiên cứu và thiếu bằng chứng, suy đoán này vẫn chưa được xác nhận, cơ sở phân tử và cơ chế bám dính mà nó tạo ra vẫn chưa rõ ràng. Trong nghiên cứu này, lần đầu tiên các nhà nghiên cứu đã xác định được một loạt gen biểu hiện cụ thể trong lớp nội bì của rễ cây lúa thông qua phân tích tin sinh học. Trong số đó, ba gen chưa được xác định trước đây có trình tự rất giống nhau và các protein mà chúng đã được mã hóa có cấu trúc không được bảo tồn ở đầu C, giàu glycine (G), alanine (A) và proline (P) và miền lectin được bảo tồn (LE) và peptide tín hiệu bài tiết (SS) ở đầu N. Các nhà nghiên cứu đặt tên cho nhóm này là GAPLESS. Những protein này hiện diện rộng rãi trong thực vật và được biểu hiện đặc biệt trong các tế bào chưa được phân hủy của nội bì. Khi các gen mã hóa ba protein GAPLESS bị loại bỏ riêng lẻ, chỉ có gen đột biến gapless1 (làm liền mạch) biểu hiện các khiếm khuyết về tăng trưởng và cân bằng nội môi ion rõ ràng như giảm chiều cao cây, giảm đẻ nhánh, năng suất cá thể thấp hơn, giảm nồng độ ion kali và tăng nồng độ ion canxi. Tuy nhiên, đột biến kép Gapless1/2 cho thấy kiểu hình tăng trưởng và cân bằng nội môi ion cực kỳ nghiêm trọng với hiệu suất dưới 3% so với kiểu hoang dã và hàm lượng ion kali chỉ bằng 10% so với kiểu hoang dã. Những kết quả này cho thấy tầm quan trọng của các protein này và dự đoán sự dư thừa chức năng của các gen này. Nghiên cứu sau đó cho thấy chức năng ngăn cản của nội bì ở rễ của đột biến đơn Gapless1 để ngăn chặn sự khuếch tán tự do của các phân tử hòa tan trong nước giảm, trong khi ở đột biến kép Gapless1/2 thậm chí còn bị suy giảm nghiêm trọng hơn. Tuy nhiên, điều tra sâu hơn cho thấy rằng sự tích lũy CS lignin của chúng không ảnh hưởng, thay vào đó CS trong đột biến kép gapless1/2 có khe hở lớn hơn và mạnh hơn. Điều này cho thấy các lỗ hổng ngăn cản cản của chúng không phải nguyên nhân do CS gây ra. Khi có sự tham gia của CS và CSD là cần thiết để thiết lập hàng rào ở tế bào nội bì nên các nhà nghiên cứu đã sử dụng kính hiển vi điện tử để quan sát cấu trúc này ở thực vật hoang dã và thực vật đột biến. Kết quả cho thấy sự tham gia của CS và CSD yếu đi ở đột biến đơn gapless1, và khoảng một nửa liên kết bị mất hoàn toàn ở đột biến kép gapless1/2 không khe hở, thậm chí liên kết còn lại bị thu hẹp nghiêm trọng. Những phát hiện này chỉ ra rằng họ protein GAPLESS cần thiết cho sự liên kết của CS và CSD và sự kết hợp tế bào rất quan trọng cho sự cân bằng dinh dưỡng và sự phát triển sinh trưởng của cây lúa. Nghiên cứu sâu hơn cho thấy rằng các protein GAPLESS được định vị cụ thể ở CS và gần với protein OsCASP1, được xác định cụ thể trong CSD. Một loạt bằng chứng thực nghiệm cho thấy protein GAPLESS có thể tương tác mạnh với OsCASP1, tạo thành phức hợp GAPLESS-OsCASP mạnh mẽ. Do đó, protein GAPLESS được gắn với CS và OsCASP và được gắn với CSD tạo thành phức hợp ổn định, CS bám với màng tế bào từ đó ngăn chặn sự khuếch tán tự do của nước và ion trong rễ. Việc làm sáng tỏ các đặc điểm phân tử và cơ chế hoạt động của GAPLESS không chỉ tăng thêm hiểu biết về chức năng của protein thành tế bào mà còn mở rộng kiến thức về cơ chế kết dính của tế bào ở sinh vật đa bào. Nghiên cứu này làm sáng tỏ bí ẩn trước đây về cơ chế phân tử làm cơ sở cho sự hình thành liên kết màng tế bào CS và xác định tầm quan trọng của liên kết chặt chẽ này đối với sự cân bằng dinh dưỡng và sự phát triển sinh trưởng của thực vật. Nó có ý nghĩa quan trọng trong việc cải thiện hiệu quả sử dụng chất dinh dưỡng khoáng và giải mã các cơ chế chịu mặn và hạn hán của thực vật, vì CS đóng một vai trò quan trọng trong việc hấp thụ có chọn lọc và phản ứng với các căng thẳng như hạn hán và nhiễm mặn của thực vật. Theo iasvn.org
Việt Nam tăng 15 bậc về Chính phủ điện tử ( 20/09/2024 )

Việt Nam tăng 15 bậc về Chính phủ điện tử...
Hỗ trợ truyền thông Cuộc thi ý tưởng dự án khởi nghiệp đổi mới sáng tạo tỉnh Tiền Giang năm 2024 ( 16/08/2024 )

Hỗ trợ truyền thông Cuộc thi ý tưởng dự án khởi nghiệp đổi mới sáng tạo tỉnh Tiền Giang năm 2024...
V/v phối hợp truyền thông về sự kiện CamaUP'24 và Cuộc thi Khởi nghiệp Cà Mau 2024 ( 14/08/2024 )
V/v phối hợp truyền thông về sự kiện CamaUP'24 và Cuộc thi Khởi nghiệp Cà Mau 2024...
Thủ tướng: “Khoa học công nghệ là con đường ngắn nhất đạt các mục tiêu” ( 17/05/2024 )

Thủ tướng: “Khoa học công nghệ là con đường ngắn nhất đạt các mục tiêu”...
✭ BÀI VIẾT LIÊN QUAN |




Truy cập hôm nay : 3
Truy cập trong 7 ngày :12
Tổng lượt truy cập : 8,919
|